
Historia de Robusta
Introducción
Las raíces de Coffea canephora, comúnmente llamada robusta[1], se remontan cientos de miles de años, al sotobosque de bosques húmedos de hoja perenne de las zonas tropicales del África subsahariana central y occidental, una región con una amplia distribución geográfica desde Guinea hasta Uganda y Angola (Dussert et al., 1999). Es una de las dos especies del género Coffea (que comprende un total de 131 especies; Davis & Rakotonasolo, 2021) que se cultivan comercialmente a escala mundial, y es reconocible visualmente por sus flores excepcionalmente grandes y su copa amplia y extendida.
A lo largo de la historia, esta especie se ha cultivado en diversas formas y ecotipos, y en regiones más allá de su lugar de origen. Hoy en día, robusta se cultiva comercialmente en unos 20 países de todo el mundo caracterizados por climas cálidos o humedad alta. La diversidad genética de robusta es enorme y, aunque los científicos llevan investigando desde la década de 1980, todavía hay mucho por descubrir sobre la especie. Sin embargo, podemos afirmar algo sobre robusta: ha evolucionado mucho para convertirse en una fuerza importante del mercado.
En las últimas décadas, el cultivo de robusta ha aumentado de manera constante y pasó del 25 % al 40 % de la producción mundial total de café desde principios de la década de 1990 (Abacusbio, 2023, pág. 13). Hasta ahora, Coffea arabica —la otra especie dominante de café comercial a nivel mundial— ha predominado en la mayor parte del mercado del café debido a las preferencias por su rendimiento de la taza. Muchos factores, incluido el aumento de la demanda de café, han provocado una expansión de la producción de robusta en los últimos años. Solo en 2023, se consumieron 177 millones de bolsas de 60 kilogramos de café en todo el mundo, lo que representa un aumento del 2,2 % con respecto al año de producción 2022 – 23 y un incremento significativo del 4,5 % con respecto al 2019 – 2020 (Organización Internacional del Café, 2024), y no se espera que la demanda disminuya.
En vista de la demanda creciente y los efectos negativos esperados del cambio climático y la limitada inversión histórica en innovación agrícola del café, un papel más importante de robusta en el mercado mundial del café es algo práctico y necesario. Sin embargo, el conocimiento sobre robusta es algo limitado entre compradores, consumidores y científicos, y aún no se ha aprovechado todo su potencial para satisfacer la demanda del mercado.
Acerca de robusta: una especie con un potencial sin explotar
Robusta tiene su origen silvestre en África subsahariana central y occidental, principalmente en el sotobosque de bosques húmedos de hoja perenne de baja altitud (50−1500 m), pero también en bosques húmedos estacionalmente secos o bosques de galería (Davis et al., 2006).
Gran parte del movimiento de robusta más allá de sus centros de origen y domesticación, así como el aumento de la popularidad de su producción a principios del siglo XX, se pueden atribuir à la propagación de la roya de la hoja de café, una enfermedad fúngica que devastó las plantas de café arábica y sigue siendo una preocupación mundial importante. Uno de los mayores beneficios de la producción de robusta es que algunas variedades poseen una resistencia natural excepcionalmente alta a las principales plagas y enfermedades que impiden una producción exitosa y rentable, incluida la roya; estas plantas a menudo pueden prosperar en condiciones adversas (Campuzano-Duque & Blair, 2022). Esta resiliencia o robustez es una de las razones por las que se suele denominar coloquialmente a Coffea canephora como robusta (nomenclatura utilizada por primera vez por Linden en 1900; Dagoon, 2005).
El crecimiento mundial de la demanda de robusta ha sido impulsado por la llegada del café soluble y la creciente popularidad de robusta en mezclas. La expansión de la producción de robusta para satisfacer esta demanda ha sido posible porque robusta puede crecer en zonas no aptas para arábica. Por ejemplo, puede producir altos rendimientos y mantener la resiliencia al estrés en rangos de temperaturas más cálidos y húmedos que arábica, que suele darse en elevaciones más bajas (entre 200 y 800 metros sobre el nivel del mar; Slipchenko, 2021). Las plantas de robusta suelen tener un mayor rendimiento, contienen mayores niveles de cafeína, menores niveles de azúcar, mayores niveles de sólidos solubles y son menos susceptibles a plagas y enfermedades dañinas (Goldemberg et al., 2015).
Muchos observadores especulan que robusta podría servir cada vez más como una alternativa a arábica a medida que el aumento de las temperaturas y la alteración de los patrones de precipitación hagan que el cultivo de arábica sea más desafiante en las próximas décadas (Bunn et al., 2015; de Aquino et al., 2022; Dinh et al., 2022; Kath et al., 2022; Kath et al., 2023). A pesar de las oportunidades que presenta robusta, enfrenta desafíos considerables propios. Una de las principales amenazas para la producción sostenible y a largo plazo de robusta de diversos orígenes son las disparidades a veces pronunciadas en la productividad[2] y rentabilidad[3] causadas por diversos factores, que incluyen los costos laborales y la mayor competencia con otros cultivos. Además, a pesar de su “robustez” frecuentemente citada, robusta sigue siendo sensible a las alteraciones ambientales. Las plantas de robusta suelen tener necesidades de precipitaciones altas, y las investigaciones recientes sugieren que la capacidad de robusta para prosperar en climas más cálidos puede haber sido sobredimensionada anteriormente; las temperaturas superiores a 20,5 °C pueden tener un impacto negativo significativo en el rendimiento de la producción (Kath et al., 2020; Tournebize et al., 2022).
Además, muchas variedades de robusta son al menos algo susceptibles a enfermedades y plagas clave, como la enfermedad de la marchitez del cafeto, la enfermedad de la ampolla roja, el barrenador del tallo, la enfermedad de la baya del café, la broca del fruto del café, los nematodos e incluso la roya de la hoja de café, entre otras (Vega et al., 2006).
Otra diferencia clave entre robusta y arábica se relaciona con el sabor y la calidad de taza (Leroy et al., 2006). El café elaborado a partir de granos de robusta suele tener menor acidez, mayor amargor y más “cuerpo” debido a su contenido de pirazina (Semmelroch & Grosch, 1995), un compuesto aromático conocido por su terrosidad. Si bien el rendimiento de la taza de robusta suele menospreciarse, cuando se manipula y procesa adecuadamente, puede servir como producto para mercados especializados (Organismo Nacional de Uganda de Desarrollo del Café, 2019). Los esfuerzos en el manejo agronómico y procesamiento posterior à la cosecha, incluido el control de calidad de los procesos de fermentación, pueden marcar una diferencia inmediata y crucial en la producción y la calidad de robusta. Sin embargo, aún queda mucho territorio por explorar en la optimización del rendimiento de robusta en el campo y de su rendimiento de la taza a escala comercial.
La historia de una especie: la historia del cultivo y la dispersión de robusta
El primer cultivo documentado de robusta comenzó alrededor de 1870 en el Congo, utilizando material genético procedente de la región del río Lomani en lo que hoy es la República Democrática del Congo (Berthaud & Charrier, 1988). Sin embargo, es probable que la producción a pequeña escala por parte de agricultores particulares y sus familias ya existiera durante décadas antes de ese momento. Un subtipo de robusta llamado “kouillou” (más tarde rebautizado “conilon” por la distorsión lingüística cuando se introdujo en Brasil) fue observado en estado silvestre por los franceses en 1880 entre Gabón y la desembocadura del río Congo, principalmente a lo largo de la región del río Kouilou-Nari. La especie fue bautizada por el botánico Louis Pierre en 1895. Pierre, que trabajaba en el Museo Nacional de Historia Natural en Francia, recibió una muestra de la planta recolectada en Gabón por el reverendo Théophile Klaine. El nombre —Coffea canephora var. Pierre ex A. Froehner— fue publicado por primera vez junto con una descripción de la especie por Froehner (1897).
Un año después, Edouard Luja fue enviado a recolectar especies con potencial económico en lo que entonces se llamaba el Congo Belga (ahora República Democrática del Congo) en preparación para la Exposición de París de 1900. Durante esta misión, Luja recogió varios miles de semillas de una “nueva” especie de café encontrada en una plantación temprana de robusta en la región (Benoit, 1968). El Congo Belga se convirtió en uno de los principales centros de diversidad, desde donde se distribuyeron líneas nuevas por todo el trópico.
A finales de siglo, la especie empezó a extenderse a otras partes del mundo. Las semillas de robusta del Congo se enviaron a Bruselas y desde allí se enviaron bajo el nombre de “robusta” a Java, Indonesia, donde fueron rápidamente aceptadas por los agricultores debido a su alta productividad y aparente resistencia à la roya de la hoja del café, especialmente porque un importante brote de la enfermedad afectó a las plantas de C. arabica en el sudeste asiático a finales del siglo XIX (Cramer, 1957). De hecho, fue en Indonesia donde se llevó a cabo por primera vez un mejoramiento genético pionero y sistemático de robusta (Ferwerda, 1948). Estos materiales se enriquecieron posteriormente con material genético procedente de Gabón y Uganda. En la misma época, otro material de robusta seleccionado de poblaciones silvestres se introdujo en zonas de Costa de Marfil, Guinea y Uganda (Charrier y Eskes, 1997).
Luego, robusta siguió extendiéndose por todo el mundo e ingresó a India a través de Java, con posteriores introducciones desde África occidental. El material seleccionado en Java se reintrodujo en África central a partir de 1910 y en la República Democrática del Congo (entonces el Congo Belga) en 1916 en el Instituto Nacional de Estudios Agronómicos del Congo Belga (INEAC), que se convirtió en centro de selección entre 1930 y 1960. Dentro de África, robusta se produjo en Madagascar, Uganda, Ghana y Costa de Marfil. Las variantes endémicas a menudo se mezclaron con las introducidas a partir de la producción comercial en otras partes del continente.
Posteriormente, robusta se introdujo en América Latina, y el grupo conilon se introdujo en Brasil en 1912 en Espírito Santo. Entre 1930 y 1935, se produjeron introducciones comerciales adicionales en América Central a través de Guatemala.
En la actualidad, solo seis países —Vietnam, Brasil, Indonesia, Uganda, India y Costa de Marfil— producen el 95 % de la variedad robusta del mundo (Abacusbio, 2023). A estas naciones les siguen Laos, Tanzania, Madagascar y Tailandia, que conforman el resto de las diez principales regiones productoras a nivel mundial. Los países de Asia y Oceanía son, en conjunto, los mayores productores de robusta, con un 60 % de la producción mundial: 41,5 millones de sacos de 60 kg al año. A esta región le sigue Sudamérica, que produce el 28 % de la variedad robusta del mundo: 19,8 millones de sacos de café en el año 2020 – 21 (Abacusbio, 2023).
Descubrimiento de la diversidad genética de robusta
El café robusta tiene un amplio alcance de diversidad genética y muchas subpoblaciones distintas. Las poblaciones silvestres son el principal pariente genético del café robusta, y el café cultivado ha cambiado relativamente poco con respecto a sus progenitores silvestres. Dentro de la reserva genética de robusta, existen muchas variaciones desconocidas, incluidos rasgos potencialmente beneficiosos relacionados con la producción y el rendimiento de la taza. En general, estas variaciones ocultas aún no han sido exploradas por productores y científicos. Profundizar la comprensión de la diversidad de robusta y su integración en los programas de mejoramiento genético es crucial para obtener ganancias genéticas continuas y a largo plazo.
Robusta también es un pariente genético de arábica (Bawin et al., 2020; Chadburn & Davis, 2017; Scalabrin et al., 2020) y tiene una relación lo suficientemente estrecha como para que los esfuerzos históricos de mejoramiento genético hayan logrado transferir cierta resistencia a las enfermedades de robusta a los cultivares de arábica (Bettencourt, 1973). La potencial transferencia de resistencia a enfermedades y plagas sigue siendo una opción para futuras iniciativas de mejoramiento genético.
Se utilizan muchos términos comunes distintos para describir a robusta en las zonas donde se cultiva. Entre estos términos se incluyen “robusta”, “conilon”, “nganda”, “koillou/quillou” y más. Estos términos son generalmente regionales y coloquiales, y no corresponden necesariamente a variedades o clones genéticamente distintos.
Lo que sí saben los científicos es que robusta es una especie diploide dividida en dos grandes grupos genéticos: guineano y congoleño. El grupo guineano —generalmente caracterizado por entrenudos estrechos, alto contenido de cafeína, bajo peso de los granos, resistencia à la sequía, ramificación secundaria y cosecha temprana— es originario de África centro-occidental. Por el contrario, el grupo congoleño —que generalmente tiene mayor resistencia à la roya, contenido medio de cafeína, alto peso de los granos, susceptibilidad à la sequía, entrenudos más grandes, crecimiento alto y cosecha tardía— es originario de África central (Herrera & Lambot, 2017). Entre estos dos grupos, el congoleño es el más extendido. Además, dentro de cada grupo hay diferentes poblaciones o subgrupos (ver la Figura 1 a continuación).
Cabe destacar que se ha habido introducciones masivas de cafetos de tipo congoleño en zonas de Costa de Marfil que albergan poblaciones silvestres endémicas de la reserva genética del guineano, lo que amenaza la integridad genética de las poblaciones silvestres de la reserva genética del guineano (Gnapi et al., 2022).
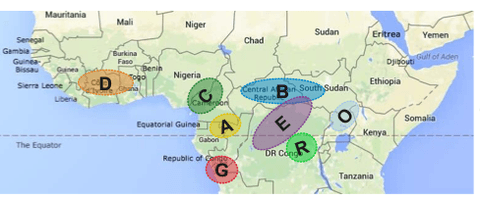
Para conservar la vasta diversidad de robusta, los bancos de genes de campo en varios países productores de África y Asia han establecido repositorios de genética de robusta (Bramel et al., 2017). A partir de la década de 1960, se llevaron a cabo actividades de recolección e intercambio de robusta coordinadas a nivel mundial desde los “centros de domesticación”. Entre las misiones de recolección importantes se incluyó una misión a Costa de Marfil, en la que ORSTROM recopiló 700 genotipos en colaboración con el Centro de Cooperación Internacional en Investigación Agrícola para el Desarrollo. Además, la especie se recolectó en Guinea, Camerún, el Congo y la República Centroafricana. A partir de estas misiones de recolección, se introdujo robusta en bancos de genes de campo de todo el mundo, entre otros, en Camerún, Costa de Marfil, Madagascar, India y Costa Rica. Dondequiera que se haya introducido el germoplasma de robusta, este ha enfrentado, y sigue enfrentando, importantes desafíos de gestión y erosión genética (Bramel et al., 2017).
Robusta en los campos de los agricultores
Debido à la necesidad de polinización cruzada de robusta (un solo clon de robusta no puede polinizar con éxito sus propias flores, como pueden hacerlo los árboles de arábica) que requiere polen de dos tipos diferentes de plantas para producir cerezas nuevas (los científicos denominan este tipo de reproducción sexual “alogamia”; Nowak, et al., 2011), los subtipos cultivados en el mismo campo suelen reproducen entre sí (Thomas, 1935). Por esta razón, es necesario que los agricultores cultiven más de un tipo de clon de robusta en sus campos para garantizar una polinización y producción de frutos exitosas. Por lo tanto, las plantaciones de robusta nunca son genéticamente uniformes. Históricamente, los agricultores de robusta tenían poco conocimiento acerca de qué variedades o subtipos estaban cultivando, aunque este conocimiento está aumentando.
La mayoría de las variedades de robusta cultivadas actualmente consisten en árboles que se originan a partir de semillas de polinización abierta (Labouisse et al., 2020) o clones multilíneas (estas variedades, que también se denominan “policlonales”, se componen de una mezcla intencional de clones genéticamente distintos; Campuzano-Duque & Blair, 2022; Montagnon et al., 2003; Berthaud & Charrier, 1998).
Sin embargo, no todos los tipos de robusta pueden crecer juntos exitosamente en un campo. La compatibilidad cruzada de los tipos está controlada genéticamente; es decir, algunas variedades son incapaces de fertilizarse entre sí (Lashermes et al., 1996; Prakash, 2018). Hasta ahora, la investigación sobre combinaciones óptimas de subtipos en producción ha sido escasa, pero una consideración clave es la floración simultánea (Silva et al., 2024).
En diferentes regiones de producción, la liberación y distribución de dichas mezclas a los agricultores se gestiona de manera diferente. Por ejemplo, en África Occidental, es habitual que los cultivadores creen variedades de semillas policlonales (es decir, varios tipos diferentes de robusta se distribuyen juntos en los mismos paquetes de semillas a los productores). En Brasil, es más común que los cultivadores creen múltiples clones únicos que luego se someten a pruebas de compatibilidad; los clones complementarios de mayor rendimiento se propagan y se ponen à la venta para los productores como plántulas propagadas de forma clonal (Depolo, et al., 2022, Surya, 2018).
El futuro de robusta
Pronto, el mundo podría enfrentar desafíos para satisfacer la creciente demanda de robusta, tal como ocurre actualmente con arábica (Abacusbio, 2023). Si bien han surgido algunas iniciativas de mejoramiento genético importantes en institutos nacionales de café en Asia, África y América Latina, aún existe una oportunidad importante para que los científicos comprendan mejor la diversidad genética de robusta y la aprovechen a través de iniciativas de mejoramiento genético modernizadas. Esto puede fortalecer la viabilidad a largo plazo de la especie para los agricultores centrándose en rasgos como el rendimiento, la resistencia a las enfermedades, la resiliencia climática y las mejoras selectivas en el rendimiento de la taza. Las colaboraciones entre científicos e instituciones pueden mejorar los servicios y las herramientas compartidos, ampliar el conocimiento y acelerar las iniciativas para desarrollar e implementar innovaciones. Abrir el camino à la innovación transformadora en el cultivo de robusta ayudará a salvaguardar el bienestar de las comunidades productoras de café y de la industria mundial del café en su conjunto.
Notas al pie
[1] A lo largo de este ensayo, y en el catálogo en general, utilizamos el término “robusta” para referirnos a toda la especie C. canephora y sus subtipos.
[2] Comparar 1,9 bolsas/ha en Costa de Marfil con 10,4 o 47,7 bolsas/ha en Uganda y Vietnam respectivamente; ver el Cuadro 1 en Abacusbio, 2023.
[3] Ver Martinez, 2023; Hasan et al., 2020.
References
Abacusbio. (2023). Opportunities for robusta variety innovation [White paper]. World Coffee Research. https://worldcoffeeresearch.org/resources/opportunities-for-robusta-variety-innovation
Bawin, Y., Ruttink, T., Staelens, A., Haegeman, A., Stoffelen, P., Mwanga, J. I. M., Roldán‐Ruiz, I., Honnay, O., & Janssens, S. B. (2020). Phylogenomic analysis clarifies the evolutionary origin of Coffea arabica. Journal of Systematics and Evolution, 59(5), 953 – 963. https://doi.org/10.1111/jse.12694
Benoit, P. L. G. (1968). Luja (Edouard Pierre). In Biographie belge d’Outremer (pp. 676 – 678). Académie Royale des Sciences d’Outré-Mer.
Berthaud, J., & Charrier, A. (1988). Use and value of genetic resources of Coffea for breeding and their long-term conservation. In Mitteilungen aus dem Institut für allgemeine Botanik in Hamburg (Vol. 23a, pp. 53 – 64). https://horizon.documentation.ird.fr/exl-doc/pleins_textes/pleins_textes_5/b_fdi_30-30/31603.pdf
Berthaud, J. (1986). Les ressources génétiques pour l’amélioration des caféiers africains diploides [Doctoral thesis, University of Paris]. Orstom. https://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers11-12/16623.pdf
Bettencourt, A. J. (1973). Considerações gerais sobre o ‘Híbrido de Timor’ (Instituto Agronômico de Campinas. Circular nº 23:20).
Botanic Gardens Conservation International, PlantSearch. (2023, March 9). https://www.bgci.org
Bramel, P., Krishnan, S., Horna, D., Lainoff, B., & Montagnon, C. (2017). Global conservation strategy for coffee genetic resources. Crop Trust & World Coffee Research. https://worldcoffeeresearch.org/resources/global-coffee-conservation-strategy
Bunn, C., Läderach, P., Ovalle Rivera, O., & Kirschke, D. (2015). A bitter cup: Climate change profile of global production of Arabica and Robusta coffee. Climatic Change, 129(1), 89 – 101. https://doi.org/10.1007/s10584-014‑1307‑0
Campuzano-Duque, L. F., & Blair, M. W. (2022). Strategies for robusta coffee (Coffea canephora) improvement as a new crop in Colombia. Agriculture, 12(10), 1576. https://doi.org/10.3390/agriculture12101576
Chadburn, H., & Davis, A. P. (2017). Coffea stenophylla. The IUCN Red List of Threatened Species. International Union for Conservation of Nature and Natural Resources. http://dx.doi.org/10.2305/IUCN.UK.2017 – 3.RLTS.T18538903A18539566.en
Cramer, P. J. S., & Wellman, F. L. (1957). A review of literature of coffee research in Indonesia. Inter-American Institute of Agricultural Sciences. https://repositorio.iica.int/handle/11324/14860
Charrier, A., & Eskes, A. B. (1997). Les caféiers. In A. Charrier et al. (Eds.), L’Amélioration des Plantes Tropicales (pp. 171 – 196). CIRAD-Orstom. https://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers09-03/010012930.pdf
Charr, J., Garavito, A., Guyeux, C., Crouzillat, D., Descombes, P., Fournier, C., Ly, S. N., Raharimalala, E. N., Rakotomalala, J., Stoffelen, P., Janssens, S., Hamon, P., & Guyot, R. (2020). Complex evolutionary history of coffees revealed by full plastid genomes and 28,800 nuclear SNP analyses, with particular emphasis on Coffea canephora (Robusta coffee). Molecular Phylogenetics and Evolution, 151, 106906. https://doi.org/10.1016/j.ympev.2020.106906
Cubry, P., Pot, D., De Bellis, F., Legnaté, H., & Leroy, T. (2008). Genetic structure of Coffea canephora Pierre species assessed by microsatellite markers [Conference presentation]. Plant and Animal Genomes XVIth Conference, San Diego, CA, United States.
Dagoon, J. D. (2005). Agriculture & Fishery Technology. Rex Bookstore, Inc.
Davis, A. P., Govaerts, R., Bridson, D. M., & Stoffelen, P. (2006). An annotated taxonomic conspectus of the genus Coffea (Rubiaceae). Botanical Journal of the Linnean Society, 152(4), 465 – 512. https://doi.org/10.1111/j.1095 – 8339.2006.00584.x
Davis, A. P., Tosh, J., Ruch, N., & Fay, M. F. (2011). Growing coffee: Psilanthus (Rubiaceae) subsumed on the basis of molecular and morphological data; implications for the size, morphology, distribution and evolutionary history of Coffea. Botanical Journal of the Linnean Society, 167(4), 357 – 377. https://doi.org/10.1111/j.1095 – 8339.2011.01177
Davis, A. P., & Rakotonasolo, F. (2021). Six new species of coffee (Coffea) from northern Madagascar. Kew Bulletin, 76(3), 497 – 511. https://doi.org/10.1007/s12225-021 – 09952‑5
Daviron, B., & Ponte, S. (2005). The coffee paradox: Global markets, commodity trade and the elusive promise of development. Zed Books.
de Aquino, S. O., Kiwuka, C., Tournebize, R., Gain, C., Marraccini, P., Mariac, C., … & Poncet, V. (2022). Adaptive potential of Coffea canephora from Uganda in response to climate change. Molecular Ecology, 31(6), 1800 – 1819. https://doi.org/10.1111/mec.16360
Depolo, R. P., Rocha, R. B., Souza, C. A. D., Santos, M. R. A. D., Espindula, M. C., & Teixeira, A. L. (2022). Expression of self-incompatibility in Coffea canephora genotypes grown in the western Amazon. Pesquisa Agropecuária Brasileira, 57. https://doi.org/10.1590/S1678-3921.pab2022.v57.03031
Dinh, T. L. A., Aires, F., & Rahn, E. (2022). Statistical analysis of the weather impact on Robusta coffee yield in Vietnam. Frontiers in Environmental Science, 10, 880. https://doi.org/10.3389/fenvs.2022.820916
Dussert, S., Lashermes, P., Anthony, F., Montagnon, C., Trouslot, P., Combes, M. C., … & Hamon, S. (1999). Le caféier, Coffea canephora. In Diversité génétique des plantes tropicales cultivées (pp. 175 – 194). https://agritrop.cirad.fr/391712/7/ID391712.pdf
Ferwerda, F. P. (1948). Coffee breeding in Java. Economic Botany, 2(3), 258 – 272. https://doi.org/10.1007/bf02859068
Froehner, A. (1897). Notizblatt des Königl. Botanischen Gartens und Museums zu Berlin, 1, 234. https://www.biodiversitylibrary.org/page/28795724#page/279/mode/1up
Gnapi, D. E., Pokou, D. N., Legnate, H., et al. (2022). Is the genetic integrity of wild Coffea canephora from Ivory Coast threatened by hybridization with introduced coffee trees from Central Africa? Euphytica, 218, 62. https://doi.org/10.1007/s10681-022 – 03004‑0
Goldemberg, D. (2019). Phenotypic and genetic characterization of the Coffea canephora collection at the University of São Paulo (USP). Revista Brasileira de Ciências Agrárias, 14(1), 70 – 75. https://doi.org/10.5039/agraria.v14n1a5088
Gonzalez, M. (2019). Coffea canephora (Robusta coffee): A review of its genetic diversity and breeding. Journal of Agronomy and Crop Science, 205(1), 1 – 17. https://doi.org/10.1111/jac.12329
Griffin, K. J., & Smith, D. B. (2015). Genetic improvement of coffee. In H. D. Schaefer & J. E. Edwards (Eds.), The chemistry of coffee (pp. 89 – 106). Springer. https://doi.org/10.1007/978 – 3‑319 – 24122-5_5
Harcourt, P. E., Ahn, S. J., & Annor, B. (2021). Genetic resources of coffee: Status and perspectives. Frontiers in Plant Science, 12, 753236. https://doi.org/10.3389/fpls.2021.753236
Hernández, E. H., Hogg, K. T., & Kreiger, N. (2019). Towards a more resilient coffee industry: How climate change impacts production. International Journal of Agricultural Sustainability, 17(1), 1 – 17. https://doi.org/10.1080/14735903.2019.1597978
Jouannic, S., & Esnault, D. (2005). Le caféier Robusta (Coffea canephora): Variétés, hybridation, production. Cirad.
Jouannic, S., Fleck, M., & Noirot, M. (2018). Genetic variation and hybridization in Coffea canephora from a production area in Central Africa. Frontiers in Plant Science, 9, 139. https://doi.org/10.3389/fpls.2018.00139
Kakuhenzire, I. S., & Ddumba, D. (2013). Coffee production in Uganda: A review of recent trends and challenges. Uganda Journal of Agricultural Sciences, 14(1), 61 – 72. https://www.ajol.info/index.php/ujas/article/view/103109
Kotsiras, A., & Varotsos, C. (2022). The genetics of coffee: A review on progress and future perspectives. Plant Biotechnology Journal, 20(1), 1 – 15. https://doi.org/10.1111/pbi.13760
Krishnan, S., & Montagnon, C. (2015). The future of coffee: A breeding perspective. In J. A. A. R. Davison & A. M. C. R. Esteves (Eds.), Coffee: Emerging trends (pp. 19 – 37). Wiley. https://doi.org/10.1002/9781118839358.ch2
Lafleur, C., & Montagnon, C. (2022). The value of coffee genetic resources: Where are we now and where do we need to go? Journal of Agricultural and Food Chemistry, 70(8), 2542 – 2554. https://doi.org/10.1021/acs.jafc.1c08378
Leigh, C., & Crouzillat, D. (2022). Genomic insights into Coffea canephora: Comparative analysis of transcriptomes and metabolomes. Molecular Plant, 15(4), 590 – 610. https://doi.org/10.1016/j.molp.2021.11.011
Lemieux, J., Mottet, A., & Olsson, A. (2021). Global challenges for coffee sustainability: Adapting to climate change and sustainable sourcing. Sustainability, 13(5), 2471. https://doi.org/10.3390/su13052471
López, C. F., & Orozco, D. C. (2022). Genomic tools for coffee genetic resources conservation: Strategies for the future. Frontiers in Plant Science, 13, 850356. https://doi.org/10.3389/fpls.2022.850356
Mekonnen, A. A., & Woldemariam, K. A. (2021). Evaluation of Coffea canephora germplasm from Uganda for resistance to coffee leaf rust and quality traits. Journal of Crop Improvement, 35(5), 699 – 711. https://doi.org/10.1080/15427528.2021.1971287
Meyer, N. (2017). Coffee in the new economy: The role of scientific innovation and technology. Agroecology and Sustainable Food Systems, 41(3), 281 – 296. https://doi.org/10.1080/21683565.2016.1251137
Musa, I., Achour, S., Poncet, V., & Lerbs, M. (2022). Coffea canephora genetic resources: Impacts on coffee production and industry. Sustainability, 14(16), 10293. https://doi.org/10.3390/su1416…
Nicolas, P., & Jaffré, T. (2017). Coffea canephora: A review of the ecological and economic significance of Robusta coffee. African Journal of Agricultural Research, 12(26), 2303 – 2313. https://doi.org/10.5897/AJAR2017.12773
Oliveira, M. A., Rodrigues, M. M., & Silva, A. G. (2022). Genetic diversity and population structure of Coffea canephora using molecular markers. Molecular Biology Reports, 49, 1017 – 1031. https://doi.org/10.1007/s11033-021 – 06943‑5
Orozco, D. C., & López, C. F. (2020). The role of coffee genetic resources in sustainable agriculture. Plant Science, 302, 110677. https://doi.org/10.1016/j.plantsci.2020.110677
Pérez, J. A., & Villegas, S. L. (2019). Climate change and coffee quality: A review of impacts and adaptation strategies. Agricultural and Forest Meteorology, 265, 56 – 64. https://doi.org/10.1016/j.agrformet.2018.11.013
Phillips, M., & Davi, K. (2021). The impact of climate change on coffee production: Implications for future research and policy. Climate Policy, 21(1), 118 – 132. https://doi.org/10.1080/14693062.2020.1790223
Ponce, D., & Ponce, J. (2020). Coffea canephora: Genetic diversity, conservation and utilization. Genetic Resources and Crop Evolution, 67(4), 889 – 902. https://doi.org/10.1007/s10722-020 – 00971‑y
Pérez, L. R., & Carrillo, A. C. (2023). Genomic and phenotypic characterization of Coffea canephora: A new perspective for coffee breeding. Plant Biotechnology Journal, 21(5), 992‑1005. https://doi.org/10.1111/pbi.13779
Santos, M. A. R. D., & D’Avila, M. A. (2021). Advances in coffee breeding in Brazil. In Plant Breeding Reviews (Vol. 45, pp. 193 – 225). Wiley. https://doi.org/10.1002/9781119677712.ch5
Segrè, A. (2019). Historical perspectives on coffee production and consumption: Socio-economic implications. Journal of Cultural Heritage Management and Sustainable Development, 9(4), 337 – 347. https://doi.org/10.1108/JCHMSD-07 – 2018-0074
Siqueira, T. L. D., & Almeida, A. C. (2021). Resilience in coffee production: Genetic diversity and adaptability in a changing climate. BMC Plant Biology, 21, 275. https://doi.org/10.1186/s12870-021 – 02920‑5
Tao, L. Y., & Geng, S. S. (2022). Coffea canephora: Genetic improvement for better resilience to climate change. Agronomy, 12(5), 1169. https://doi.org/10.3390/agronomy12051169
Teixeira, A. M., & Gomes, L. F. (2019). Perspectives on coffee breeding: Genetic diversity and sustainable production. Plant Breeding, 138(5), 526 – 533. https://doi.org/10.1111/pbr.12696
Turgut, O., & Güler, S. (2021). Genetic diversity and climate resilience in Coffea canephora: Strategies for sustainable coffee production. Journal of Agricultural and Food Chemistry, 69(8), 2348 – 2362. https://doi.org/10.1021/acs.jafc.0c05870
Valerio, A., & Mangini, G. (2020). Conservation of genetic resources in Coffea canephora: Current status and future strategies. Frontiers in Plant Science, 11, 1 – 12. https://doi.org/10.3389/fpls.2020.00001
Wang, M., & Tian, J. (2022). Strategies for enhancing the resilience of Coffea canephora to climate change: A review. Agricultural Sciences, 13(4), 391 – 405. https://doi.org/10.4236/as.2022.134032
Wong, T. H., & Thong, H. C. (2022). Genomics and breeding of Coffea canephora: Advances and prospects. Journal of Agricultural Science, 14(5), 25 – 38. https://doi.org/10.5539/jas.v14n5p25
Zhao, M., & Liu, Y. (2021). Assessing the genetic diversity of Coffea canephora using SSR markers: Implications for conservation and breeding. Genetics and Molecular Research, 20(4), gmr18555. https://doi.org/10.4238/gmr18555
World Coffee Research
WCR es un programa de investigación y desarrollo colaborativo sin fines de lucro 501 (c)(5) de la industria mundial del café para cultivar, proteger y mejorar los suministros de café de calidad mientras se mejoran los medios de vida de las familias que lo producen.
¿Es usted un productor de café?
Hemos creado una versión especial de nuestro Catálogo de variedades de café especialmente para productores. Disponible en inglés y español.