
History of Robusta
Introduction
The roots of Coffea canephora, commonly called robusta[1], trace back hundreds of thousands of years to the humid, understory evergreen forests in tropical areas of central and western sub-Saharan Africa — a region with expansive geographic distribution from Guinea to Uganda to Angola (Dussert et al., 1999). It is one of the two species in the Coffea genus (which comprises a total of 131 species; Davis & Rakotonasolo, 2021) that are commercially cultivated on a global scale, and it is visually recognizable by its uniquely large blossoms and wide, spreading canopy.
Throughout history, this species has been grown in numerous forms and ecotypes, and in regions beyond its birthplace. Today, robusta is cultivated commercially in about 20 countries worldwide, characterized by warm climates and/or high humidity. Robusta’s genetic diversity is vast, and while scientists have been researching this since the 1980s, there is still much to uncover about the species. Nonetheless, one thing about robusta is certain: it has dramatically evolved into a major market force.
In recent decades, robusta cultivation has steadily increased, rising from 25% to 40% of total global coffee production since the early 1990s (Abacusbio, 2023, p. 13). Until now, Coffea arabica—the other globally dominant commercial coffee species — has held sway over most of the coffee market due to preferences for its cup quality. Many factors, including increasing demand for coffee, have led to expansions in robusta production in recent years. In 2023 alone, 177 million 60-kilogram bags of coffee were consumed worldwide — a 2.2% increase from the 2022 – 23 production year and a striking 4.5% jump from 2019 – 2020 (International Coffee Organization, 2024) — and demand is not expected to slow down.
With soaring demand at center stage, alongside the expected negative effects of climate change and limited historical investment in coffee agricultural innovation, an expanded role for robusta in the global coffee market is both practical and necessary. However, understanding of robusta is somewhat limited among buyers, consumers, and scientists alike, and its full potential in meeting market demand has yet to be realized.
About robusta — a species of untapped potential
Robusta has its wild origins in central and western sub-Saharan Africa, mainly in the humid understory of low-elevation evergreen forests (50 – 1500 m), but sometimes in seasonally dry humid forests or gallery forests (Davis et al., 2006).
Much of the movement of robusta beyond its centers of origin and domestication, as well as the increase in the popularity of its production during the early 1900s, can be attributed to the spread of coffee leaf rust — a fungal disease that ravaged arabica coffee plants and remains a significant global concern. One of the greatest benefits of robusta production is that some varieties possess an exceptionally high natural resistance to major pests and diseases that impede successful and profitable production, including rust; these plants can often thrive under harsh conditions (Campuzano-Duque & Blair, 2022). This resilience or robustness is one reason why Coffea canephora is often colloquially referred to as robusta (nomenclature that was first used by Linden in 1900; Dagoon, 2005).
Global growth in demand for robusta has been driven by the advent of soluble coffee and the increasing popularity of robusta in blends. The expansion of robusta production to meet this demand has been possible because robusta can grow in areas unsuitable for arabica. For example, it can produce high crop yields and maintain stress resilience in hotter, more humid temperature ranges than arabica, which typically occur at lower elevations (between 200 – 800 meters above sea level; Slipchenko, 2021). Robusta plants typically have greater crop yields, contain higher levels of caffeine, lower levels of sugar, higher levels of soluble solids, and are less susceptible to damaging pests and diseases (Goldemberg et al., 2015).
Many observers speculate that robusta may increasingly serve as an alternative to arabica as rising temperatures and altered precipitation patterns make arabica cultivation more challenging in the coming decades (Bunn et al., 2015; de Aquino et al., 2022; Dinh et al., 2022; Kath et al., 2022; Kath et al., 2023). Despite the opportunities presented by robusta, it faces considerable challenges of its own. One primary threat to long-term, sustainable robusta production from diverse origins is the sometimes steep disparities in productivity[2] and profitability[3], caused by numerous factors, including labor costs and increased competition from other crops. Additionally, despite its oft-cited “robustness,” robusta is still sensitive to environmental disturbances. Robusta plants generally have high precipitation needs, and recent research suggests that robusta’s ability to thrive in hotter climates may have been previously overstated; temperatures over 20.5 degrees Celsius can have significantly negative impact on yields (Kath et al., 2020; Tournebize et al., 2022).
Moreover, many robusta varieties are at least somewhat susceptible to key diseases and pests, such as coffee wilt disease, red blister disease, stem borer, coffee berry disease, coffee berry borer, nematodes, and even coffee leaf rust, among others (Vega et al., 2006).
Another key difference between robusta and arabica pertains to taste and cup quality (Leroy et al., 2006). Coffee brewed from robusta beans is often lower in acidity, higher in bitterness, and more “full-bodied” due to its pyrazine content (Semmelroch & Grosch, 1995), an aromatic compound known for its earthiness. While the cup quality of robusta is often disparaged, when handled and processed properly, it can serve as a product for specialty markets (Uganda Coffee Development Authority, 2019). Agronomic and post-harvest processing efforts, including quality control of fermentation processes, can make an immediate and crucial difference in robusta production and quality. However, there remains significant unexplored territory in optimizing robusta’s performance in the field and its cup quality on a commercial scale.
The story of a species: The history of robusta’s cultivation and dispersal
The first documented cultivation of robusta began around 1870 in Congo, using genetic material from the Lomani River region in what is now the Democratic Republic of Congo (Berthaud & Charrier, 1988). However, it is likely that small-scale production by individual farmers and their families had been ongoing for decades prior to this point. A subtype of robusta called “koillou” (later renamed “conilon” through linguistic distortion when it was introduced to Brazil) was observed in the wild by the French in 1880 between Gabon and the mouth of the Congo River, mainly along the Kouilou-Niari River region. The species was named by the botanist Louis Pierre in 1895. Pierre, who worked at the Muséum National d’Histoire Naturelle in France, received a sample of the plant collected in Gabon by the Reverend Théophile Klaine. The name—Coffea canephora var. Pierre ex A. Froehner — was first published along with a description of the species by Froehner (1897).
One year later, Edouard Luja was sent to collect species with economic potential in what was then called Belgian Congo (now the Democratic Republic of Congo) in preparation for the 1900 Paris Exposition. During this mission, Luja collected several thousand seeds from a “new” coffee species found in an early robusta plantation in the region (Benoit, 1968). Belgian Congo became one of the principal centers of diversity, from which new lines were distributed throughout the tropics.
At the turn of the century, the species began to spread to other parts of the world. Robusta seeds from Congo were sent to Brussels, and from there they were dispatched under the name “robusta” to Java, Indonesia, where they were quickly accepted by farmers due to their high productivity and apparent resistance to coffee leaf rust — especially as a major outbreak of the disease affected C. arabica plants in Southeast Asia in the late 1800s (Cramer, 1957). In fact, it was in Indonesia that pioneering, systematic robusta breeding was first conducted (Ferwerda, 1948). These materials were later enriched with genetic material from Gabon and Uganda. Around the same time, other robusta material selected from wild populations was introduced to areas of Ivory Coast, Guinea, and Uganda (Charrier and Eskes, 1997).
From here, robusta continued to spread around the world, entering India via Java, with later introductions from West Africa. Material selected in Java was reintroduced to Central Africa from 1910 onward and to the Democratic Republic of Congo (then Belgian Congo) in 1916 at the Institut National pour l’Étude Agronomique du Congo (INEAC), which became the center for selection from 1930 to 1960. Within Africa, robusta was produced in Madagascar, Uganda, Ghana, and Ivory Coast. Endemic variants often intermingled with those introduced from commercial production in other parts of the continent.
Robusta was later introduced to Latin America, with the conilon group introduced in Brazil in 1912 to Espiritu Santo. Additional commercial introductions occurred in Central America via Guatemala between 1930 and 1935.
Today, just six countries — Vietnam, Brazil, Indonesia, Uganda, India, and Côte d’Ivoire — produce 95% of the world’s robusta (Abacusbio, 2023). Laos, Tanzania, Madagascar, and Thailand follow these nations in production, making up the remainder of the top ten producing regions worldwide. Countries in Asia and Oceania are collectively the largest producers of robusta, generating 60% of the world’s output at 41.5 million 60 kg bags annually. This region is followed by South America, which produces 28% of the world’s robusta, generating 19.8 million bags of coffee in the 2020 – 21 year (Abacusbio, 2023).
Discovering robusta’s genetic diversity
Robusta coffee has a wide scope of genetic diversity, with many distinct sub-populations. Wild populations are the primary genetic relatives of robusta coffee, and cultivated coffee has changed relatively little from its wild progenitors. Many unknown variations, including potentially beneficial traits related to production and cup quality, exist within the robusta gene pool. By and large, these hidden variations have yet to be explored and utilized by scientists and breeders. Deepening the understanding of robusta diversity and its integration into breeding programs is crucial for continuous and long-term genetic gains.
Robusta is also a genetic relative of arabica (Bawin et al., 2020; Chadburn & Davis, 2017; Scalabrin et al., 2020) and is closely related enough that historical breeding efforts have successfully transferred some disease resistance from robusta into arabica cultivars (Bettencourt, 1973). Potential disease and pest resistance transference remains possible for future breeding efforts.
Many different common terms are used to describe robusta in the areas where it is grown. These terms include “robusta,” “conilon,” “nganda,” “koillou/quillou,” and others. These terms are generally regional and colloquial, and they do not necessarily correspond to specific genetically distinct varieties or clones.
What scientists do know is that robusta is a diploid species divided into two broad genetic groups: Guinean and Congolese. The Guinean group — which is generally characterized by narrow internodes, high caffeine content, low bean weight, drought resistance, secondary branching, and early harvest — originated in central-west Africa. In contrast, the Congolese group — which typically has higher rust resistance, medium caffeine content, high bean weight, drought susceptibility, larger internodes, tall growth, and late harvesting — originated in central Africa (Herrera & Lambot, 2017). Among these two groups, the Congolese is the most widespread. Additionally, within each group, there are different populations or subgroups (see Figure 1 below).
It is noteworthy to highlight that there have been massive introductions of Congolese-type trees into areas of Côte d’Ivoire that are home to wild endemic populations from the Guinean gene pool, which threaten the genetic integrity of wild populations from the Guinean gene pool (Gnapi et al., 2022).
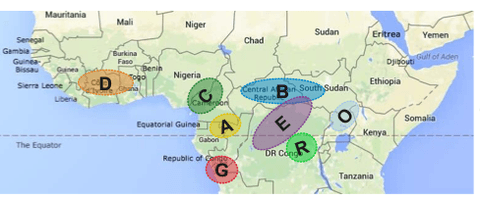
To conserve robusta’s vast diversity, field gene banks in several producing countries in Africa and Asia have established repositories of robusta genetics (Bramel et al., 2017). Targeted, globally coordinated robusta collection and exchange were conducted starting in the 1960s from “centers of domestication.” Important collecting missions included a mission to Côte d’Ivoire, which collected 700 wild genotypes by ORSTROM in collaboration with the Centre de Coopération Internationale en Recherché Agronomique pour le Développement. Additionally, the species has been collected in Guinea, Cameroon, the Congo, and the Central African Republic. From these collecting missions, robusta was introduced to field genebanks around the world, including in Cameroon, Ivory Coast, Madagascar, India, and Costa Rica, among others. Wherever robusta germplasm has been introduced, it has faced — and continues to face — significant management and genetic erosion challenges (Bramel et al., 2017).
Robusta in farmers’ fields
Because robusta necessarily cross-pollinates — a single robusta clone cannot successfully pollinate its own flowers, as arabica trees can do — but requires pollen from two different types of plants to produce new cherries (scientists call this “allogamous”; Nowak, 2011) — subtypes grown in the same field typically interbreed (Thomas, 1935). For this reason, it is necessary for farmers to grow more than one type of robusta clone in their fields to ensure successful pollination and fruit production. Robusta plantations are therefore never genetically uniform. Historically, robusta farmers had little awareness of which varieties or subtypes they were growing, although this awareness is increasing.
Most currently cultivated robusta consists either of trees originating from open-pollinated seeds (Labouisse et al., 2020) or multi-line clones (these varieties, which are also called “polyclonal,” are comprised of an intentional mix of genetically distinct clones; Campuzano-Duque & Blair, 2022; Montagnon et al., 2003; Berthaud & Charrier, 1998).
However, not all robusta types can successfully grow together in a field. The cross-compatibility of types is genetically controlled; in other words, some varieties are unable to fertilize one another (Lashermes et al., 1996; Prakash, 2018). So far, research on optimal combinations of subtypes in production has been scarce, but one key consideration is simultaneous flowering (Silva et al., 2024).
In different production regions, the release and distribution of such mixes to farmers is handled differently. For example, in West Africa, it is common for breeders to create polyclonal seed varieties (i.e., multiple different types of robusta are distributed together in the same seed packets to farmers). In Brazil, it is more common for breeders to create multiple unique clones that are then tested for compatibility; the highest-performing complementary clones are then propagated and released to farmers as clonally propagated seedlings (Depolo et al., 2022; Prakash, 2018).
The future of robusta
The world may soon face challenges in keeping up with the rising demand for robusta, just as it currently does for arabica (Abacusbio, 2023). While some strong breeding initiatives have emerged at national coffee institutes across Asia, Africa, and Latin America, there remains a significant opportunity for scientists to better understand robusta’s genetic diversity and leverage it through modernized breeding initiatives. This can strengthen the species’ long-term viability for farmers by focusing on traits such as yield, disease resistance, climate resilience, and selective improvements in cup quality. Collaborations between scientists and institutions can enhance shared services and tools, expand knowledge, and accelerate efforts to develop and deploy innovations. Clearing a path for transformative innovation in robusta will help safeguard the well-being of coffee farming communities and the global coffee industry as a whole.
Footnotes
[1] Throughout this essay and the catalog generally, we use this term“robusta” to refer to the entire C. canephora species and all its subtypes.
[2] Compare 1.9 bags/ha in Côte d’Ivoire to 10.4 or 47.7 bags/ha in Uganda and Vietnam respectively; see Table 1 in Abacusbio, 2023.
[3] See Martinez, 2023; Hasan et al., 2020.
References
Abacusbio. (2023). Opportunities for robusta variety innovation [White paper]. World Coffee Research. https://worldcoffeeresearch.org/resources/opportunities-for-robusta-variety-innovation
Bawin, Y., Ruttink, T., Staelens, A., Haegeman, A., Stoffelen, P., Mwanga, J. I. M., Roldán‐Ruiz, I., Honnay, O., & Janssens, S. B. (2020). Phylogenomic analysis clarifies the evolutionary origin of Coffea arabica. Journal of Systematics and Evolution, 59(5), 953 – 963. https://doi.org/10.1111/jse.12694
Benoit, P. L. G. (1968). Luja (Edouard Pierre). In Biographie belge d’Outremer (pp. 676 – 678). Académie Royale des Sciences d’Outré-Mer.
Berthaud, J., & Charrier, A. (1988). Use and value of genetic resources of Coffea for breeding and their long-term conservation. In Mitteilungen aus dem Institut für allgemeine Botanik in Hamburg (Vol. 23a, pp. 53 – 64). https://horizon.documentation.ird.fr/exl-doc/pleins_textes/pleins_textes_5/b_fdi_30-30/31603.pdf
Berthaud, J. (1986). Les ressources génétiques pour l’amélioration des caféiers africains diploides [Doctoral thesis, University of Paris]. Orstom. https://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers11-12/16623.pdf
Bettencourt, A. J. (1973). Considerações gerais sobre o ‘Híbrido de Timor’ (Instituto Agronômico de Campinas. Circular nº 23:20).
Botanic Gardens Conservation International, PlantSearch. (2023, March 9). https://www.bgci.org
Bramel, P., Krishnan, S., Horna, D., Lainoff, B., & Montagnon, C. (2017). Global conservation strategy for coffee genetic resources. Crop Trust & World Coffee Research. https://worldcoffeeresearch.org/resources/global-coffee-conservation-strategy
Bunn, C., Läderach, P., Ovalle Rivera, O., & Kirschke, D. (2015). A bitter cup: Climate change profile of global production of Arabica and Robusta coffee. Climatic Change, 129(1), 89 – 101. https://doi.org/10.1007/s10584…
Campuzano-Duque, L. F., & Blair, M. W. (2022). Strategies for robusta coffee (Coffea canephora) improvement as a new crop in Colombia. Agriculture, 12(10), 1576. https://doi.org/10.3390/agriculture12101576
Chadburn, H., & Davis, A. P. (2017). Coffea stenophylla. The IUCN Red List of Threatened Species. International Union for Conservation of Nature and Natural Resources. http://dx.doi.org/10.2305/IUCN…
Cramer, P. J. S., & Wellman, F. L. (1957). A review of literature of coffee research in Indonesia. Inter-American Institute of Agricultural Sciences. https://repositorio.iica.int/handle/11324/14860
Charrier, A., & Eskes, A. B. (1997). Les caféiers. In A. Charrier et al. (Eds.), L’Amélioration des Plantes Tropicales (pp. 171 – 196). CIRAD-Orstom. https://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers09-03/010012930.pdf
Charr, J., Garavito, A., Guyeux, C., Crouzillat, D., Descombes, P., Fournier, C., Ly, S. N., Raharimalala, E. N., Rakotomalala, J., Stoffelen, P., Janssens, S., Hamon, P., & Guyot, R. (2020). Complex evolutionary history of coffees revealed by full plastid genomes and 28,800 nuclear SNP analyses, with particular emphasis on Coffea canephora (Robusta coffee). Molecular Phylogenetics and Evolution, 151, 106906. https://doi.org/10.1016/j.ympev.2020.106906
Cubry, P., Pot, D., De Bellis, F., Legnaté, H., & Leroy, T. (2008). Genetic structure of Coffea canephora Pierre species assessed by microsatellite markers [Conference presentation]. Plant and Animal Genomes XVIth Conference, San Diego, CA, United States.
Dagoon, J. D. (2005). Agriculture & Fishery Technology. Rex Bookstore, Inc.
Davis, A. P., Govaerts, R., Bridson, D. M., & Stoffelen, P. (2006). An annotated taxonomic conspectus of the genus Coffea (Rubiaceae). Botanical Journal of the Linnean Society, 152(4), 465 – 512. https://doi.org/10.1111/j.1095 – 8339.2006.00584.x
Davis, A. P., Tosh, J., Ruch, N., & Fay, M. F. (2011). Growing coffee: Psilanthus (Rubiaceae) subsumed on the basis of molecular and morphological data; implications for the size, morphology, distribution and evolutionary history of Coffea. Botanical Journal of the Linnean Society, 167(4), 357 – 377. https://doi.org/10.1111/j.1095 – 8339.2011.01177
Davis, A. P., & Rakotonasolo, F. (2021). Six new species of coffee (Coffea) from northern Madagascar. Kew Bulletin, 76(3), 497 – 511. https://doi.org/10.1007/s12225-021 – 09952‑5
Daviron, B., & Ponte, S. (2005). The coffee paradox: Global markets, commodity trade and the elusive promise of development. Zed Books.
de Aquino, S. O., Kiwuka, C., Tournebize, R., Gain, C., Marraccini, P., Mariac, C., … & Poncet, V. (2022). Adaptive potential of Coffea canephora from Uganda in response to climate change. Molecular Ecology, 31(6), 1800 – 1819. https://doi.org/10.1111/mec.16360
Depolo, R. P., Rocha, R. B., Souza, C. A. D., Santos, M. R. A. D., Espindula, M. C., & Teixeira, A. L. (2022). Expression of self-incompatibility in Coffea canephora genotypes grown in the western Amazon. Pesquisa Agropecuária Brasileira, 57. https://doi.org/10.1590/S1678-3921.pab2022.v57.03031
Dinh, T. L. A., Aires, F., & Rahn, E. (2022). Statistical analysis of the weather impact on Robusta coffee yield in Vietnam. Frontiers in Environmental Science, 10, 880. https://doi.org/10.3389/fenvs.2022.820916
Dussert, S., Lashermes, P., Anthony, F., Montagnon, C., Trouslot, P., Combes, M. C., … & Hamon, S. (1999). Le caféier, Coffea canephora. In Diversité génétique des plantes tropicales cultivées (pp. 175 – 194). https://agritrop.cirad.fr/391712/7/ID391712.pdf
Ferwerda, F. P. (1948). Coffee breeding in Java. Economic Botany, 2(3), 258 – 272. https://doi.org/10.1007/bf02859068
Froehner, A. (1897). Notizblatt des Königl. Botanischen Gartens und Museums zu Berlin, 1, 234. https://www.biodiversitylibrary.org/page/28795724#page/279/mode/1up
Gnapi, D. E., Pokou, D. N., Legnate, H., et al. (2022). Is the genetic integrity of wild Coffea canephora from Ivory Coast threatened by hybridization with introduced coffee trees from Central Africa? Euphytica, 218, 62. https://doi.org/10.1007/s10681-022 – 03004‑0
Goldemberg, D. (2019). Phenotypic and genetic characterization of the Coffea canephora collection at the University of São Paulo (USP). Revista Brasileira de Ciências Agrárias, 14(1), 70 – 75. https://doi.org/10.5039/agrari…
Gonzalez, M. (2019). Coffea canephora (Robusta coffee): A review of its genetic diversity and breeding. Journal of Agronomy and Crop Science, 205(1), 1 – 17. https://doi.org/10.1111/jac.12…
Griffin, K. J., & Smith, D. B. (2015). Genetic improvement of coffee. In H. D. Schaefer & J. E. Edwards (Eds.), The chemistry of coffee (pp. 89 – 106). Springer. https://doi.org/10.1007/978 – 3-…
Harcourt, P. E., Ahn, S. J., & Annor, B. (2021). Genetic resources of coffee: Status and perspectives. Frontiers in Plant Science, 12, 753236. https://doi.org/10.3389/fpls.2…
Hernández, E. H., Hogg, K. T., & Kreiger, N. (2019). Towards a more resilient coffee industry: How climate change impacts production. International Journal of Agricultural Sustainability, 17(1), 1 – 17. https://doi.org/10.1080/147359…
Jouannic, S., & Esnault, D. (2005). Le caféier Robusta (Coffea canephora): Variétés, hybridation, production. Cirad.
Jouannic, S., Fleck, M., & Noirot, M. (2018). Genetic variation and hybridization in Coffea canephora from a production area in Central Africa. Frontiers in Plant Science, 9, 139. https://doi.org/10.3389/fpls.2…
Kakuhenzire, I. S., & Ddumba, D. (2013). Coffee production in Uganda: A review of recent trends and challenges. Uganda Journal of Agricultural Sciences, 14(1), 61 – 72. https://www.ajol.info/index.ph…
Kotsiras, A., & Varotsos, C. (2022). The genetics of coffee: A review on progress and future perspectives. Plant Biotechnology Journal, 20(1), 1 – 15. https://doi.org/10.1111/pbi.13…
Krishnan, S., & Montagnon, C. (2015). The future of coffee: A breeding perspective. In J. A. A. R. Davison & A. M. C. R. Esteves (Eds.), Coffee: Emerging trends (pp. 19 – 37). Wiley. https://doi.org/10.1002/978111…
Lafleur, C., & Montagnon, C. (2022). The value of coffee genetic resources: Where are we now and where do we need to go? Journal of Agricultural and Food Chemistry, 70(8), 2542 – 2554. https://doi.org/10.1021/acs.ja…
Leigh, C., & Crouzillat, D. (2022). Genomic insights into Coffea canephora: Comparative analysis of transcriptomes and metabolomes. Molecular Plant, 15(4), 590 – 610. https://doi.org/10.1016/j.molp…
Lemieux, J., Mottet, A., & Olsson, A. (2021). Global challenges for coffee sustainability: Adapting to climate change and sustainable sourcing. Sustainability, 13(5), 2471. https://doi.org/10.3390/su1305…
López, C. F., & Orozco, D. C. (2022). Genomic tools for coffee genetic resources conservation: Strategies for the future. Frontiers in Plant Science, 13, 850356. https://doi.org/10.3389/fpls.2…
Mekonnen, A. A., & Woldemariam, K. A. (2021). Evaluation of Coffea canephora germplasm from Uganda for resistance to coffee leaf rust and quality traits. Journal of Crop Improvement, 35(5), 699 – 711. https://doi.org/10.1080/154275…
Meyer, N. (2017). Coffee in the new economy: The role of scientific innovation and technology. Agroecology and Sustainable Food Systems, 41(3), 281 – 296. https://doi.org/10.1080/216835…
Musa, I., Achour, S., Poncet, V., & Lerbs, M. (2022). Coffea canephora genetic resources: Impacts on coffee production and industry. Sustainability, 14(16), 10293. https://doi.org/10.3390/su1416…
Nicolas, P., & Jaffré, T. (2017). Coffea canephora: A review of the ecological and economic significance of Robusta coffee. African Journal of Agricultural Research, 12(26), 2303 – 2313. https://doi.org/10.5897/AJAR20…
Oliveira, M. A., Rodrigues, M. M., & Silva, A. G. (2022). Genetic diversity and population structure of Coffea canephora using molecular markers. Molecular Biology Reports, 49, 1017 – 1031. https://doi.org/10.1007/s11033…
Orozco, D. C., & López, C. F. (2020). The role of coffee genetic resources in sustainable agriculture. Plant Science, 302, 110677. https://doi.org/10.1016/j.plan…
Pérez, J. A., & Villegas, S. L. (2019). Climate change and coffee quality: A review of impacts and adaptation strategies. Agricultural and Forest Meteorology, 265, 56 – 64. https://doi.org/10.1016/j.agrf…
Phillips, M., & Davi, K. (2021). The impact of climate change on coffee production: Implications for future research and policy. Climate Policy, 21(1), 118 – 132. https://doi.org/10.1080/146930…
Ponce, D., & Ponce, J. (2020). Coffea canephora: Genetic diversity, conservation and utilization. Genetic Resources and Crop Evolution, 67(4), 889 – 902. https://doi.org/10.1007/s10722…
Pérez, L. R., & Carrillo, A. C. (2023). Genomic and phenotypic characterization of Coffea canephora: A new perspective for coffee breeding. Plant Biotechnology Journal, 21(5), 992‑1005. https://doi.org/10.1111/pbi.13…
Santos, M. A. R. D., & D’Avila, M. A. (2021). Advances in coffee breeding in Brazil. In Plant Breeding Reviews (Vol. 45, pp. 193 – 225). Wiley. https://doi.org/10.1002/978111…
Segrè, A. (2019). Historical perspectives on coffee production and consumption: Socio-economic implications. Journal of Cultural Heritage Management and Sustainable Development, 9(4), 337 – 347. https://doi.org/10.1108/JCHMSD…
Siqueira, T. L. D., & Almeida, A. C. (2021). Resilience in coffee production: Genetic diversity and adaptability in a changing climate. BMC Plant Biology, 21, 275. https://doi.org/10.1186/s12870…
Tao, L. Y., & Geng, S. S. (2022). Coffea canephora: Genetic improvement for better resilience to climate change. Agronomy, 12(5), 1169. https://doi.org/10.3390/agrono…
Teixeira, A. M., & Gomes, L. F. (2019). Perspectives on coffee breeding: Genetic diversity and sustainable production. Plant Breeding, 138(5), 526 – 533. https://doi.org/10.1111/pbr.12…
Turgut, O., & Güler, S. (2021). Genetic diversity and climate resilience in Coffea canephora: Strategies for sustainable coffee production. Journal of Agricultural and Food Chemistry, 69(8), 2348 – 2362. https://doi.org/10.1021/acs.ja…
Valerio, A., & Mangini, G. (2020). Conservation of genetic resources in Coffea canephora: Current status and future strategies. Frontiers in Plant Science, 11, 1 – 12. https://doi.org/10.3389/fpls.2…
Wang, M., & Tian, J. (2022). Strategies for enhancing the resilience of Coffea canephora to climate change: A review. Agricultural Sciences, 13(4), 391 – 405. https://doi.org/10.4236/as.202…
Wong, T. H., & Thong, H. C. (2022). Genomics and breeding of Coffea canephora: Advances and prospects. Journal of Agricultural Science, 14(5), 25 – 38. https://doi.org/10.5539/jas.v1…
Zhao, M., & Liu, Y. (2021). Assessing the genetic diversity of Coffea canephora using SSR markers: Implications for conservation and breeding. Genetics and Molecular Research, 20(4), gmr18555. https://doi.org/10.4238/gmr185…
World Coffee Research
World Coffee Research is a 501 (c)(5) non-profit, collaborative research and development program of the global coffee industry to grow, protect, and enhance supplies of quality coffee while improving the livelihoods of the families who produce it.
Are you a coffee farmer?
We've created a printable version of our coffee variety catalog specifically for farmers. Available in English and Spanish.